
Within the last few years, my hometown has seen a fortunate increase in both microbreweries and local distilleries. At one of my new favorite places to purchase craft beers I came across a beer that inspired this blog post. The Scottish brewed Fraoch Heather Ale boasts right on the bottle that the recipe originated in Scotland, 4000 years ago. “Leann Fraoch”, Gaelic for heather ale, is a light amber ale made from malted wheat, barley and hops (the usual suspects for beer) with bog myrtle (an herb native to Scotland) and heather, the purple shrub so indicative of the British Isles. While the heather ale brewed in Scotland today by Williams Bros Brewing Company is no doubt a reconstruction (using modern ingredients for a modern palette with modern fermentation processes), there is in fact evidence of a fermented drink containing heather dating back to 2ooo B.C. in Scotland. Analysis of Neolithic pot sherds from the Isle of Rhum revealed the residue of mashed cereal straw, cereal-type pollen, meadowsweet, heather, and royal fern that researchers have interpreted as an ancient brew of Northern Europe (Gregory et al. 2005; Nelson, 2005).
This investigation into the Neolithic origins of modern Scottish beer sparked other questions for me concerning drinking in the past. When in the past did we begin exploiting the process of fermentation? What lines of evidence contribute to our understanding of past fermentation practices and the recreation of these recipes? And finally, is it possible to identify an alcoholic from skeletal remains of such an individual? Throughout this post, I will explore these questions as I delve into the archaeological, historical, and biochemical literature concerning fermentation.

Humans & Alcohol
Humans have been enjoying/lamenting/over-indulging in the fruits of their fermented labor, namely drink, for thousands of years across the world. Whether you are a heavy drinker or completely abstain, it is difficult to deny that alcohol has been intertwined with human culture, politics, and social life since its inception. We will discuss that and how we identify alcohol in the archaeological record in a moment, but for now we have to back up a bit. When considering a topic using an anthropological lens, we must first consider the non-humans.
For those of you who may not know, the etymology of anthropology is from the Greek anthropos (man/human/humanity) and –logy (branch of learning/study of). Therefore, the very word anthropology presupposes that there is indeed a study of humans – that we are unique enough to warrant a branch of social science devoted to study. As a result, one of the important initial questions to ask when examining human behavior is, is this behavior uniquely human?
Enjoying the breakdown of sugar by yeast is an activity in which both humans and non-humans (animals, insects, etc) engage – sometimes to excess. Through its bright color and sweet flavor, fruit attracts animals large and small in a symbiotic relationships that feeds the animals and disperses seeds of the flowering plants. Birds have been known to overindulge on overripe fruit and fall of their perch or even die from ethanol poisoning. Elephants in Africa have broken into moonshine headquarters and in a drunken rampage, destroyed property and even killed people. In perhaps my favorite example, Charles Darwin took advantage of a shared love of booze when he laid out bowls of beer at night and in the morning was able to collect drunk baboons as docile specimens (McGovern 2009). It is also highly probable that our ancestors enjoyed a fruit cocktail from time to time. Studies examining both dental micro and macro morphology suggest that a wide variety of hominid species most likely consumed fruit as part of their diet in Africa (Ward et al. 1999; Wood & Richmond, 2000). Indeed, our closest living relative, the chimpanzee, has a diet comprised of 71% fruit, and favors those that are ripe (Tweheyo et al. 2004). In fact, for some anthropologists, alcohol helped make us human.
In Robert Dudley’s The Drunken Monkey: Why We Drink and Abuse Alcohol, he expands on a hypothesis originally proposed in 2000 that the human attraction to alcohol consumption is biological – that at one point it provided us an evolutionary advantage. If many of our ancestors were primarily fruit eaters, they may have been enjoying moderate amounts of alcohol – perhaps even remembering and exploiting the fact that overly ripe fruit produces a sweet cocktail. While the ability to smell and identify alcohol molecules is well documented in fruit flies (Kaun et al. 2016; Versace et al. 2016), apparently it has been assumed that such olfactory abilities were non-existent in primates (Dudley 2014). However, behavioral and physiological tests of this assumption over the last decade have displayed that a variety of primate species can in fact smell and taste organic compounds like ethanol that are the result of fermentation (Dominy 2004; Laska & Seibt 2002).
Two predictions from evolutionary medicine can help explain why it is not out of the question that we have an evolutionary relationship with alcohol. Firstly, Dudley (2014) explains that human abuse of alcohol may be related to the same mechanism that drives obesity. Simply put, in the past when our ancestors were foraging and even hunting, calories could be scarce, so food sources were always heavily exploited when they were available. Perhaps our response to alcohol is no different. In regard to ethanol or other alcohol molecules, “If natural selection has acted on human ancestors to associate this molecule with nutritional gain, then its psychoactive features may simply promote evolved and once advantageous behaviors that encourage its rapid consumption” (Dudley 2014:67). Secondly, through natural selection animals can also acquire the ability to take advantage of low-level toxic compounds with regular exposure (Dudley 2000; 2014). Indeed, daily moderate consumption of alcohol has been shown to have health benefits including the overall reduction of cardiovascular risk and mortality (which is why you see articles all over the internet about how you should have that daily glass of wine). The “drunken monkey” is still a hypothesis that needs more in depth testing, especially of how humans, primates, other mammals, and birds seek out and experience alcohol via fruit fermentation in the wild. However, it offers an intriguing possible explanation for our preoccupation with fermentation, both in the past and today.

The evidence for the importance, and potentially advantageous, alcohol consumption is apparent in the historical record as well once humans began fermenting their own products. From hieroglyphic inscriptions, we know that the ancient Egyptians thought quite highly of beer. This fermented beverage supplemented the diets of individuals across socioeconomic classes, served as an offering to the gods, and was even buried with deceased individuals to bring the party to the afterlife (Samuel 1996). In the Middle Ages, The Regimen Sanitatis Salernitanum (try saying that five times fast) was a document created by doctors for English royalty and from 1200-1800 was a popular guide to health in Europe (Laskow 2016). The document says a lot about wine, a little about beer, but barely mentions water. The attraction of beer and wine in these cultures may have been so prevalent partly because fermented beverages served as a reliable source of potable water. The process of fermentation kills much of the bacteria that can be lurking in unclean water. So before the understanding of germs and the utilization of widespread water treatment, beverages like beer and wine were a safe source of hydration.
The relationship between humans and alcohol is also highly relevant to our socio-cultural identity. “Fermented beverages are at the center of religions all around the world. [Alcohol] makes us who we are in a lot of ways,” says Patrick McGovern in an interview with the Smithsonian. Eating and drinking are most often social activities, so what and how we drink is a marker of identity and establishes social group boundaries that exist both within and between cultures (Nelson 2005). And now that we’ve established the importance of alcohol – to non-humans and humans alike, we can discuss how we identify its use and production in the past.
Making Our Own
While fermentation is a natural process that many other animals also enjoy, distillation is a human invention. So when in the past did we start brewing and distilling? First, a note on the interpretation of archaeological evidence before we continue. Archaeology is dedicated to the examination and analysis of past human activity. This activity is interpreted from the archaeological record – the physical evidence we find from the past. That recored can be biased since the materials used affects the preservation potential of artifacts, so that some activity may be “invisible” to the archaeological eye. Therefore, it is paramount to remember that absence of evidence is not evidence of absence.
It is impossible to discuss the archaeology of alcoholic beverages without mentioning Patrick McGovern, currently the Scientific Director of the Biomolecular Archaeology Project for Cuisine, Fermented Beverages and Health at the University of Pennsylvania Museum. He is widely regarded as the “Beer Archaeologist”, an expert on identifying fermentation in the past. Not only is he testing for evidence of what and how ancients were fermenting, but has also partnered with several brewers to recreate such recipes. His research, which began in the Middle East identifying wine from residue analysis, has taken him around the world as archaeologists seek to identify the presence, use and importance of alcohol through time. To identify the presence of alcohol in the past, McGovern and other archaeologists most often test organic residues from inside ceramic jars for the presence of trace molecules indicative of certain types of alcohol production or the DNA of yeast.
The oldest known evidence for the production of alcohol is from China around 7000 BCE during the Neolithic Period (McGovern et al. 2004). The Neolithic Period, as McGovern (2009) explains, is ripe with possibilities for evidence of distillation – if only because the earliest known pottery dates to this time period. It is not outside the realm of possibility that humans were producing fermented beverages prior to this period, or experimenting with the process, but they may have been doing so in such a way that is essentially invisible to us.
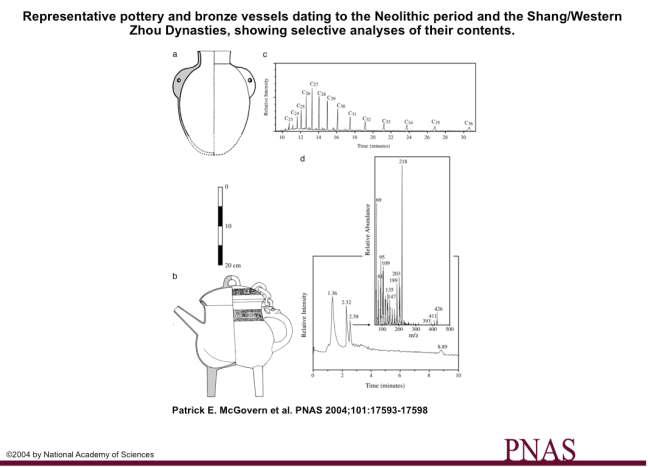
As McGovern (2009) describes in his most recent book, Uncorking the Past, a team of Chinese archaeologists invited him to join them in analyzing pot sherds they suspected once held alcohol. Jiahu, located in the Henan province of China, is a Neolithic village that dates to the early Neolithic with occupation as early as 7000 B.C. (radiocarbon) and holds evidence for the earliest musical instruments and domesticated rice in Northern China (McGovern et al. 2004). The pot sherds were subjected to a variety of tests to determine their chemical composition. The Jiahu pottery samples yielded FT-IR and HRLC matches to modern rice and rice wine, beeswax, grape tannins, and herbal constituents, while the carbon isotope analysis showed C13 values consistent with C3 plants – like rice or grapes (McGovern et al. 2004). The Chinese archaeologists and McGovern interpreted these finds as a mixed fermented beverage made of rice, honey, and fruit. However, the sequence of n-alkanes (a hydrocarbon molecule) McGovern and colleagues (2004) interpreted as beeswax in their analysis (see image below) is also characteristic with petroleum (Evershed 2008). Obviously in this instance it is more likely the compound in jar originated from the presence of beeswax but this ambiguity is what makes recreating exact recipes difficult.
Another method by which to identify a fermented beverage is to check for the presence of the fermentor, or the yeast within the mixture. In modern processes, the primary yeast used in fermentation is Saccharomyces cerevisiae, present during bread making, winemaking and brewing. And it appears that we have been utilizing this yeast for fermentation for at least 5000 years. In Egypt, archaeologists identified the presence of S. cerevisiae from residue within a jar dating to 3000 B.C. (radiocarbon), recovered from the tomb of King Scorpion I – one of the first kings of Egypt (Cavalieri et al. 2002). The DNA was extracted from residue within the jar and then amplified with lab equipment not previously exposed to modern S. cerevisiae to prevent contamination. When compared, the sequence of the extracted DNA most closely resembles the DNA sequence of modern S. cerevisiae (Cavalieri et al. 2002). An analysis of the different types of S. cerevisiae strains used in different fermentation practices, revealed that beer strains were far removed from those used for wine indicating that beer did not originate from wine technology as previously assumed (Legras et al. 2007).
Beer making, as McGovern (2009) points out, requires an extra step since grains do not naturally host yeast so it must be added to covert the starch into sugar. At Godin Tepe, a site in western Iran that acted as a trading post in ancient Mesopotamia, McGovern helped identify some of the earliest beer technology, dating to 3500-3100 B.C. Calcium oxalate is a compound also known as beerstone since it is known to be present during the processing and storing of barley beer (McGovern 2009). At Godin Tepe, beerstone was identified using a simple spot test along with the presence of six-row domesticated barley, leading researchers to believe that they had discovered the storage area, or possible brewing site, of an ancient beer (McGovern 2009). And while wine technology may have been utilized first, there is evidence from analyzing strains of S. cerevisiae that we were enjoying a brew before we were breaking bread (Legras et al. 2007).
Alcohol in our Bones?
Palaeopathology is the study of disease and pathology in the past as it is observable in skeletal assemblages (Waldron, 2009). This practice relies on differential diagnosis, since some diseases affect the bones in very similar ways. The diagnosis of a specific disease may be impossible depending on the evidence from the bones available and, if possible, a range of possible ailments may be presented (Waldron, 2009). Additionally, some diseases/conditions, like the plague, kill too quickly to ever leave evidence of their existence on bone (at least at the macro level). That being said, is it possible to identify a chronic alcoholic based on skeletal analysis?
A meta-analysis by Gaddini and colleagues (2016) examined studies that assessed the effects of alcohol on bone in adult humans. Light to moderate alcohol consumption (less than or equal to 3 drinks a week) can actually increase bone mineral density (BMD) since alcohol slows the overall rate of bone remodeling (Gaddini et al. 2016). Such a conclusion, however, should be deemed tentative as the authors point out that few studies have examined this issue. In general, high consumption of alcohol is associated with a lower BMD, but again, the authors urge caution as the range of studies they examined included no to high alteration of BMD with heavy drinking due to factors like age, duration, and pattern of alcohol consumption (Gaddini et al. 2016). However, there are other diseases that affect BMD and therefore the likelihood of bone fracture, namely osteoporosis (Waldron, 2009). Therefore, identifying alcoholics from BMD changes among adult skeletal populations would be almost impossible.

Another way to consider alcohol’s affect on the human skeleton is to examine possible hard tissue consequences of alcohol on the fetus. The most frequent Alcohol Related Birth Defects (ARBD) include microcephaly (significantly smaller head), synostosis or fusion of the radius/ulna and carpals, minor abnormalities of the joints in the hands and feet, and/or delayed growth (Shuler & Schroeder 2013). In an effort to study evidence for such alcohol related defects in the skeleton, Shuler and Schroeder

(2013) analyzed a skeletal population excavated in the late 1990s from Newton Plantation (ca. 1660-1820) in Barbados where there is evidence of consumption of possible lead containing rum by inhabitants. Out of the 45 skeletal individuals identified, two (N48 and N49) displayed evidence of congenital defects, though N48 was an adult male whose anomalies are more likely the result of lead poisoning (Shuler & Schroeder 2013).

In specimen N49, an adolescent aged 9-11 years old, Shuler & Schroeder (2013) identified multiple fusions of the cervical vertebrae that they argue differs enough from another condition (Klippel Feil Syndrome) to be malformations considered diagnostic of ARBD. The fusion rates for the 2nd and 3rd cervical vertebrae (seen above) are especially more common in Fetal Alcohol Syndrome than they are in KFS . The mandibular changes of N49 (seen directly above) are also more consistent with in utero or early childhood ethanol exposure (Schuler & Schroeder 2013).
Although the results of possible ARBD from Schuler and Schroeder (2013) are intriguing, the nature of bone’s response to many different diseases or conditions in a similar manner complicates our initial question. Due to this equifinality, positive identification of fetal/early childhood exposure to ethanol or adult alcoholism is an improbable diagnosis on its own. It is possible, given certain archaeological/cultural contexts, however, that it may be listed as a possible cause of low BMD or skeletal malformation in a differential diagnosis.
References
Cavalieri, D., McGovern, P. E., Hartl, D. L., Mortimer, R., & Polsinelli, M. (n.d.). Evidence for S. cerevisiae Fermentation in Ancient Wine. Journal of Molecular Evolution, 57(1), S226–S232. https://doi.org/10.1007/s00239-003-0031-2
Dominy, N. J. (2004). Fruits, Fingers, and Fermentation: The Sensory Cues Available to Foraging Primates. Integrative and Comparative Biology, 44(4), 295–303. https://doi.org/10.1093/icb/44.4.295
Dudley, R. (2000). Evolutionary Origins of Human Alcoholism in Primate Frugivory. The Quarterly Review of Biology, 75(1), 3–15.
Dudley, R. (2014). The Drunken Monkey: Why We Drink and Abuse Alcohol. University of California Press.
Evershed, R. P. (2008). Organic Residue Analysis in Archaeology: The Archaeological Biomarker Revolution*. Archaeometry, 50(6), 895–924. https://doi.org/10.1111/j.1475-4754.2008.00446.x
Gaddini, G. W., Turner, R. T., Grant, K. A., & Iwaniec, U. T. (2016). Alcohol: A Simple Nutrient with Complex Actions on Bone in the Adult Skeleton. Alcoholism: Clinical and Experimental Research, 40(4), 657–671. https://doi.org/10.1111/acer.13000
Gregory, R. A., Murphy, E. M., Church, M. J., Edwards, K. J., Guttmann, E. B., & Simpson, D. D. A. (2005). Archaeological evidence for the first Mesolithic occupation of the Western Isles of Scotland. The Holocene, 15(7), 944–950. https://doi.org/10.1191/0959683605hl868ft
Kaun, K. R., Azanchi, R., Maung, Z., Hirsh, J., & Heberlein, U. (2011). A Drosophila model for alcohol reward. Nature Neuroscience, 14(5), 612–619. https://doi.org/10.1038/nn.2805
Laska, M., & Seibt, A. (2002). Olfactory sensitivity for aliphatic alcohols in squirrel monkeys and pigtail macaques. Journal of Experimental Biology, 205(11), 1633–1643.
Laskow, S. (400AD, 19:00). I Tried a Medieval Diet, And I Didn’t Even Get That Drunk. Retrieved October 16, 2016, from http://www.atlasobscura.com/articles/i-tried-a-medieval-diet-and-i-didnt-even-get-that-drunk
Legras, J.-L., Merdinoglu, D., Cornuet, J.-M., & Karst, F. (2007). Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Molecular Ecology, 16(10), 2091–2102. https://doi.org/10.1111/j.1365-294X.2007.03266.x
McGovern, P. E. (2009). Uncorking the Past: The Quest for Wine, Beer, and Other Alcoholic Beverages. University of California Press.
McGovern, P. E., Zhang, J., Tang, J., Zhang, Z., Hall, G. R., Moreau, R. A., … Wang, C. (2004). Fermented beverages of pre- and proto-historic China. Proceedings of the National Academy of Sciences of the United States of America, 101(51), 17593–17598. https://doi.org/10.1073/pnas.0407921102
Nelson, M. (2005). The Barbarian’s Beverage: A History of Beer in Ancient Europe. Routledge.
Shuler, K. A., & Schroeder, H. (2013). Evaluating Alcohol Related Birth Defects in the past: Skeletal and biochemical evidence from a colonial rum producing community in Barbados, West Indies. International Journal of Paleopathology, 3(4), 235–242. https://doi.org/10.1016/j.ijpp.2013.08.005
Sicard, D., & Legras, J.-L. (2011). Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. Comptes Rendus Biologies, 334(3), 229–236. https://doi.org/10.1016/j.crvi.2010.12.016
Tweheyo, M., Lye, K. A., & Weladji, R. B. (2004). Chimpanzee diet and habitat selection in the Budongo Forest Reserve, Uganda. Forest Ecology and Management, 188(1–3), 267–278. https://doi.org/10.1016/j.foreco.2003.07.028
Versace, E., Eriksson, A., Rocchi, F., Castellan, I., Sgadò, P., & Haase, A. (2016). Physiological and behavioral responses in Drosophila melanogaster to odorants present at different plant maturation stages. Physiology & Behavior, 163, 322–331. https://doi.org/10.1016/j.physbeh.2016.05.027

Fascinating writing on alcohol & human acts! Thanks for jnywllectual perspectives & facts! Enjoyed a lot!! — your fan, Soo-Yeon from hometown,Columbia,MO
LikeLike
[…] you enjoyed my first post about the archaeology of beer and wine you may be interested to know that the Field Museum, […]
LikeLike
[…] (for a broader explanation of differential diagnosis in paleopathology, see my previous post on The Archaeology of Fermentation). As seen in the figure above, increased porosity of several bones in the skull is often sited as a […]
LikeLike
Howdy! I could have sworn I’ve visited your blog before but after going through some of the articles I realized it’s new to me. Anyhow, I’m certainly delighted I stumbled upon it and I’ll be book-marking it and checking back often!
LikeLike